Новости мира науки Выяснение отношений между птицами разных видов имеет далеко идущие последствия (08.06.2010)
 Детальный анализ пространственного распределения птиц, гнездящихся в Дании, показал, что оно в значительной мере формируется в результате их непосредственных взаимодействий. Птицы одного вида, хотя и отстаивают свою гнездовую территорию, часто селятся по соседству, а вот представители разных, но экологически близких видов расходятся в пространстве. Происходит это не за счет выбора данными видами разных типов местообитаний, а за счет избегания (или прямого вытеснения) одних видов другими. Поразительно, что выявленный эффект сказывается на распределении видов в пространстве, которое по площади в тысячи и даже в десятки тысяч раз превышает территорию, охраняемую отдельной гнездящейся парой.
Детальный анализ пространственного распределения птиц, гнездящихся в Дании, показал, что оно в значительной мере формируется в результате их непосредственных взаимодействий. Птицы одного вида, хотя и отстаивают свою гнездовую территорию, часто селятся по соседству, а вот представители разных, но экологически близких видов расходятся в пространстве. Происходит это не за счет выбора данными видами разных типов местообитаний, а за счет избегания (или прямого вытеснения) одних видов другими. Поразительно, что выявленный эффект сказывается на распределении видов в пространстве, которое по площади в тысячи и даже в десятки тысяч раз превышает территорию, охраняемую отдельной гнездящейся парой.
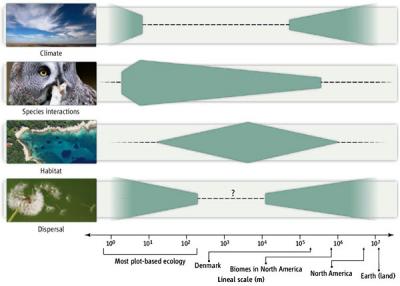
Если законы Ньютона с равным успехом объясняют движение планет и бильярдных шаров, то законы экологии нередко оказываются зависящими от масштаба, то есть работают только в определенном диапазоне расстояний и на определенных отрезках времени. Очевидно, к примеру, что участок земли размером 10 × 10 м2 представляет собой совершенно разное пространство с «точки зрения» бактерии, дождевого червя, полёвки или слона. И дело не только в том, что разное их число может прокормиться и даже попросту поместиться на таком участке. Не менее важна и разная степень гетерогенности для них одного и того же пространства, разное число местообитаний, которые они могут там найти.
Если ограничиться одной группой организмов и посмотреть, какие факторы определяют пространственное распределение отдельных видов, то выяснится, что они окажутся разными в зависимости от масштаба рассмотрения. Так, распределение птиц в пределах материков и биомов (крупных природных зон) будет определяться климатом и возможностями расселения. В пределах биома (например, зоны широколиственных лесов Европы) распределение будет зависеть в первую очередь от выбора теми или иными видами конкретных местообитаний (биотопов), например участков высокоствольного леса, опушек, зарослей кустарников и т. п. Наконец, на пространственное распределение может влиять и прямое взаимодействие («выяснение отношений») между птицами как одного, так и разных видов. Хорошо известно, что многие птицы активно защищают свою территорию (где находится гнездо и где они добывают корм для птенцов) от вторжения чужаков, причем чужаками могут быть особи как своего, так и других видов.
 Впрочем, до недавнего времени роль межвидовой конкуренции, как фактора, определяющего распределение птиц, более или менее обстоятельно изучалась только на примере островных фаун. Так, Джаред Даймонд (Jared M. Diamond), обследуя острова архипелага Бисмарка (к северо-востоку от Новой Гвинеи), показал (Diamond, 1975), что виды одного рода, как правило, обитают на разных островах, демонстрируя «шахматное распределение» (клетка может быть свободной или занята только одной фигурой — одним из видов). Работы Даймонда вызвали большой резонанс (см.: Assembly rules). Некоторые критики полагали, что размер выборки (число островов и видов птиц) недостаточно велик, чтобы делать категорические выводы о ведущей роли конкуренции в распределении разных видов по разным островам. Однако, как показали математические модели, вероятность того, что наблюдаемое распределение является чистой случайностью (а взаимодействие видов ни при чём), оказалась крайне низкой.
Впрочем, до недавнего времени роль межвидовой конкуренции, как фактора, определяющего распределение птиц, более или менее обстоятельно изучалась только на примере островных фаун. Так, Джаред Даймонд (Jared M. Diamond), обследуя острова архипелага Бисмарка (к северо-востоку от Новой Гвинеи), показал (Diamond, 1975), что виды одного рода, как правило, обитают на разных островах, демонстрируя «шахматное распределение» (клетка может быть свободной или занята только одной фигурой — одним из видов). Работы Даймонда вызвали большой резонанс (см.: Assembly rules). Некоторые критики полагали, что размер выборки (число островов и видов птиц) недостаточно велик, чтобы делать категорические выводы о ведущей роли конкуренции в распределении разных видов по разным островам. Однако, как показали математические модели, вероятность того, что наблюдаемое распределение является чистой случайностью (а взаимодействие видов ни при чём), оказалась крайне низкой.
 Но даже соглашаясь с тем, что конкуренция — действительно важный фактор, регулирующий распределение экологически близких видов по разным островам, исследователи вовсе не были уверены в том, что подобное явление наблюдается на материке, где нет такой изоляции друг от друга отдельных участков пригодных местообитаний. Более того — местообитания, в той или иной степени подходящие для какой-то группы птиц, могут непрерывно тянуться и занимать обширное пространство. Что же касается межвидовых взаимодействий птиц, то считалось, что если они и проявляются, то лишь на небольших расстояниях, сопоставимых по величине с размерами территории, охраняемой отдельной парой (для мелких певчих птиц это десятки и сотни метров).
Но даже соглашаясь с тем, что конкуренция — действительно важный фактор, регулирующий распределение экологически близких видов по разным островам, исследователи вовсе не были уверены в том, что подобное явление наблюдается на материке, где нет такой изоляции друг от друга отдельных участков пригодных местообитаний. Более того — местообитания, в той или иной степени подходящие для какой-то группы птиц, могут непрерывно тянуться и занимать обширное пространство. Что же касается межвидовых взаимодействий птиц, то считалось, что если они и проявляются, то лишь на небольших расстояниях, сопоставимых по величине с размерами территории, охраняемой отдельной парой (для мелких певчих птиц это десятки и сотни метров).
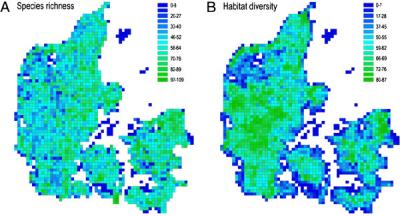
 Ясность в этот вопрос была внесена совсем недавно, в ходе исследования Николаса Готелли (Nicolas J. Gotelli), Гари Грейвса (Gary R. Graves) и Карстена Рабека (Carsten Rahbek). По их данным, эффект, оказываемый межвидовыми взаимодействиями на пространственное распределение птиц, может сказываться в весьма крупных масштабах — на территориях, площадь которых на 3–4 порядка превышает площадь индивидуальных охраняемых участков. К такому выводу они пришли, анализируя очень подробные сведения о распределении гнездящихся птиц (197 видов) на территории Дании. Исходные данные собирали 750 наблюдателей в течение 1993–1996 гг. Для учета распределения птиц и характеристики основных биотопов (местообитаний) вся территория Дании (за исключением удаленных и небольших островов) была разбита сеткой квадратов с размерами отдельной ячейки 5 × 5 км2 (всего 2003 ячейки). Площадь такой ячейки примерно соответствует охотничьей территории филина, одного из самых крупных хищников в составе охваченной орнитофауны. В другом варианте анализа пространственного распределения птиц использовали данные по ячейкам 10 × 10 км2 (всего 620 ячеек).
Ясность в этот вопрос была внесена совсем недавно, в ходе исследования Николаса Готелли (Nicolas J. Gotelli), Гари Грейвса (Gary R. Graves) и Карстена Рабека (Carsten Rahbek). По их данным, эффект, оказываемый межвидовыми взаимодействиями на пространственное распределение птиц, может сказываться в весьма крупных масштабах — на территориях, площадь которых на 3–4 порядка превышает площадь индивидуальных охраняемых участков. К такому выводу они пришли, анализируя очень подробные сведения о распределении гнездящихся птиц (197 видов) на территории Дании. Исходные данные собирали 750 наблюдателей в течение 1993–1996 гг. Для учета распределения птиц и характеристики основных биотопов (местообитаний) вся территория Дании (за исключением удаленных и небольших островов) была разбита сеткой квадратов с размерами отдельной ячейки 5 × 5 км2 (всего 2003 ячейки). Площадь такой ячейки примерно соответствует охотничьей территории филина, одного из самых крупных хищников в составе охваченной орнитофауны. В другом варианте анализа пространственного распределения птиц использовали данные по ячейкам 10 × 10 км2 (всего 620 ячеек).
Поскольку подавляющее большинство птиц Дании (более 95%) совершают дальние миграции на зимовку, а молодые особи оседлых видов также легко перемещаются на довольно значительные расстояния, предположение о том, что расселение есть фактор, ограничивающий распределение, можно было отбросить. Также, ввиду небольших размеров страны и ее ровного низменного рельефа, можно было не рассматривать и предположение о влиянии климата. Остаются два фактора: 1) выбор птицами подходящего местообитания или 2) чисто поведенческие реакции особей одного вида на присутствие поблизости особей другого вида — избегание или активная агрессия. Очевидно, что факторы эти не являются взаимоисключающими и что разграничить их очень непросто. Тем не менее авторы обсуждаемой работы попытались это сделать.
 Для решения поставленной задачи сначала выделили 12 разных типов местообитаний (морские открытые воды, пресные водоемы, болота, поля, пастбища, листопадный лес, хвойный лес и т. п.). На суше разные местообитания оказались представленными сравнительно небольшими фрагментами, разбросанными по всей территории. В результате даже в одну ячейку 5 × 5 км2 попадал целый ряд местообитаний. Чтобы оценить их разнообразие, для каждой ячейки подсчитывали специальный индекс, учитывающий представленность всех местообитаний. Затем исследователи выделили целый ряд экологических групп (гильдий) птиц, различающихся прежде всего характером пищи и способом ее добывания. Среди этих гильдий — нырковые утки, настоящие утки, дятлы, кормящиеся в кронах мелкие насекомоядные, кормящиеся на открытой местности мелкие насекомоядные, зерноядные воробьиные, дрозды и т. п. (всего 24 гильдии). Кроме того, в пределах некоторых гильдий были выделены группы близкородственных видов.
Для решения поставленной задачи сначала выделили 12 разных типов местообитаний (морские открытые воды, пресные водоемы, болота, поля, пастбища, листопадный лес, хвойный лес и т. п.). На суше разные местообитания оказались представленными сравнительно небольшими фрагментами, разбросанными по всей территории. В результате даже в одну ячейку 5 × 5 км2 попадал целый ряд местообитаний. Чтобы оценить их разнообразие, для каждой ячейки подсчитывали специальный индекс, учитывающий представленность всех местообитаний. Затем исследователи выделили целый ряд экологических групп (гильдий) птиц, различающихся прежде всего характером пищи и способом ее добывания. Среди этих гильдий — нырковые утки, настоящие утки, дятлы, кормящиеся в кронах мелкие насекомоядные, кормящиеся на открытой местности мелкие насекомоядные, зерноядные воробьиные, дрозды и т. п. (всего 24 гильдии). Кроме того, в пределах некоторых гильдий были выделены группы близкородственных видов.
Сопоставление данных по пространственному распределению всех видов и принадлежности их к той или иной гильдии анализировали с помощью моделей (матриц), отражающих разные нуль-гипотезы о связи распространения видов с определенными местообитаниями. Поскольку в реальном распределении организмов в пространстве всегда существует элемент стохастичности (конкретный вид может отсутствовать в определенной ячейке из-за случайных обстоятельств), без построения подобных моделей корректно решить поставленную задачу просто нельзя. Суть проблемы авторы сформулировали в двух вопросах:
1) Демонстрируют ли виды одной гильдии (в другом варианте — близкородственные виды) неслучайные паттерны пространственного распределения — агрегацию (совместно встречаются чаще, чем встречались бы чисто случайно) или сегрегацию (виды расходятся в пространстве)?
2) Может ли неслучайная в разных масштабах картина распределения птиц быть объяснена наличием и выбором подходящих местообитаний?
Результаты работы показали, что распределение птиц неслучайно и что в некоторых случаях местообитание играет ключевую роль (к примеру, настоящие утки встречаются только на пресных водоемах и в болотах). Однако в случае обитателей суши, особенно мелких певчих птиц, известных хорошо выраженным территориальным поведением, авторы обнаружили четкое расхождение видов одной гильдии в пространстве, и это расхождение не могло быть объяснено приуроченностью видов к разным местообитаниям. Более того, размеры участков, занимаемых разными экологически близкими видами, существенно превосходили охраняемые территории отдельных особей (разница площадей — на 3–4 порядка величин).
 Какой механизм может сформировать такую крупномасштабную пространственную структуру? Ответить на этот вопрос не так просто. Авторы обращают внимание на то, что большинство видов — это мигранты. Весной они возвращаются на еще никем не занятые места и стремятся в них сразу обосноваться. При этом самцы того же вида селятся не слишком далеко от других, уже занявших территорию, самцов того же вида. В результате представители одного вида могут заселить целиком определенный изолированный фрагмент ландшафта (например, участок леса среди полей). Прилетевшие чуть позже особи другого вида скорее поселятся на незанятом никем участке, чем будут вступать в агрессивные взаимодействия с уже обосновавшимися особями других видов. Таким образом, некоторая комбинация внутривидового (конспецифического) притяжения и межвидового отталкивания может, по мысли авторов, привести к крупномасштабному пространственному разделению близких видов.
Какой механизм может сформировать такую крупномасштабную пространственную структуру? Ответить на этот вопрос не так просто. Авторы обращают внимание на то, что большинство видов — это мигранты. Весной они возвращаются на еще никем не занятые места и стремятся в них сразу обосноваться. При этом самцы того же вида селятся не слишком далеко от других, уже занявших территорию, самцов того же вида. В результате представители одного вида могут заселить целиком определенный изолированный фрагмент ландшафта (например, участок леса среди полей). Прилетевшие чуть позже особи другого вида скорее поселятся на незанятом никем участке, чем будут вступать в агрессивные взаимодействия с уже обосновавшимися особями других видов. Таким образом, некоторая комбинация внутривидового (конспецифического) притяжения и межвидового отталкивания может, по мысли авторов, привести к крупномасштабному пространственному разделению близких видов.
Получается, что межвидовая конкуренция (точнее — поведение, позволяющее эту конкуренцию избежать) приводит к тому, что обитающие на материке экологически близкие виды птиц расходятся пространственно почти так же, как расходились бы они, заселяя разные острова.
Алексей Гиляров
Элементы



